

Introduction
草本层植被是温带森林中维管植物物种多样性最高的地方,在森林生态系统中发挥着关键的作用。它可以作为一个良好的生态指标,其物种构成是众多环境驱动因素和时空资源梯度之间复杂相互作用的结果。确定群落组成的生态决定因素已成为更好地了解森林植被将如何应对全球环境变化带来的挑战的关键一步,这些环境变化包括二氧化碳水平增加、营养沉积导致富营养化或酸化的养分沉积以及干扰制度的改变。
温带森林生态系统目前覆盖了全球森林面积的约16%。它们涵盖了各种各样的森林类型,其林下落叶层中居住的植物物种具有独特的生活史策略,表现为适应当地环境条件的功能特征。森林林下植物物种在生长和生活型、叶片寿命、物候期、克隆繁殖的流行率、演替状态和种子传播机制方面存在差异。在成熟林分中,封闭的乔木层冠层通过限制森林地面的光可获得性来诱导强烈的过滤作用。物种组成和多样性以及草本层群落的盖度可能会受到其他环境压力的影响,如养分或水分供应不足。同样,高海拔地区的冷胁迫有利于具有显著保守性的物种,如较小的叶片和较矮的植株。在资源有限的条件下生存、生长和繁殖可以通过广泛的生态策略来实现,从长寿生物(树木)到一年生草本物种。
植物功能特征与环境因素密切相关。物种特征可以告诉我们形态、物候和生理适应的领先维度,使物种能够在森林草本层中生存下来。例如,许多典型的森林物种已经进化出在落叶覆盖层树冠关闭之前完成其生命周期的具体战略(例如早春短暂植物)。其他森林专家(主要是演替后期、耐荫的多年生草本植物,局限于封闭的森林内部,很少在其他生境中找到,如森林边缘或树冠空隙)在本季度晚些时候开始生长和开花。这种时间生态位分化对森林林下植被物种共存具有重要意义。森林植物可以耐受弱光环境,并表现出独特的功能特征,例如更高的比叶面积、更大的种子和各种储存或克隆器官。此外,混合阔叶落叶林中的生境专家已证明主要在高海拔和更冷、更干燥的地点茁壮成长,并且第一次开花的年龄晚于多面手。它们的种子直径更小,叶片干物质含量更低,平均冠层高度更低,比叶面积更高。根据Grime的CSR理论,演替后期森林林下落叶层的特征是耐受胁迫或竞争物种,而干扰增加了社区水平CSR特征的原始成分。
科学问题:(1)哪些环境变量对草本层的植物区系组成影响最显著;(2)物种的生活史特征和群落CSR特征与环境变量的关系。利用分布在斯洛文尼亚各地的系统采样植被数据,我们假设草本层植被的区系和功能组成与不同的环境因素有关
Materials and methods
研究区域和植被采样:这项研究包括50个不同的森林地点。研究地表现出非常多样化的自然条件。选择的林地在地质基岩、气候、土壤类型、树种组成、林分结构等非生物和生物因素方面存在显著差异。研究区域包括从200米-1500米的森林。研究区的特点是气候梯度很大。研究地在地理和生态上处于中欧和东南欧的过渡地带,位于地中海、阿尔卑斯山和大陆气候体系的交汇处,气候变化很大,可分为六个植物地理区:高山、前阿尔卑斯、迪纳里期、迪纳里期、次地中海和亚潘尼期,物种组成差异较大。所描述的天然林类型与多种形式的次生林和其他森林群落(通常以同质结构的同龄林为特征)交织在一起,这是管理的结果,即树种组成适度和相关的非生物(土壤)条件。// 在这项研究中,我们选择了制定了监测计划的林地,39个地块属于第一级计划,11个地块是为密集监测方案(第二级)设计的。一级地块系统地分布在斯洛文尼亚境内16千米*16千米的网格中。在11个不同地点设立了二级地块,代表斯洛文尼亚森林类型的异质性。我们收集了50个面积为400平方米的监测样地的植物群落和环境数据。乔木层(高度为5米的木本植物)、灌木层(高度大于0.5米但小于5米的木本植物)和草本层(高度为0.5米的所有草本植物和木本植物)中的所有维管束植物都被记录下来,遵循地面植被采样的统一ICP协议。每种植物的丰度是根据巴克曼方法的改进进行估计的。

环境数据:对于每个研究样地,我们收集了土壤属性、林分特征、气候参数和其他环境变量的数据。根据国际比较方案方法,对每个监测区的土壤进行了采样。采集不同土层的土壤样品,区分有机土层(Of、Ol、OH)和矿物层(0~5 cm、5~10 cm、10~20 cm、20~40 cm和40~80 cm)。对不同层次的土壤样本测量了下列生态相关变量(表1):PH值(以CaCl2测量)、土壤有机碳(SOC,g/kg)、全氮含量(g/kg)、粘土比例(来自土壤质地分析)和土壤水分含量(%)。然后对各层的值进行平均,以获得单个地块级别的值。还考虑了乔木层特征:乔木层物种丰富度、阔叶层比例和乔木层遮荫能力(SCA),以加权平均值的形式计算,使用树种相对丰富度和遮荫产值(基于叶面积指数进行评估)。低的SCA值表示相对稀疏的树冠,而高的SCA值表示浓密的树冠投下较深的阴影。两个气候参数,年平均温度(MAT)和年平均降水量(MAP)。环境矩阵还包含关于地块高度、热负荷指数(Hi)的数据,Hi表示考虑到局部坡向和坡度的太阳辐射接收的相对热量,木质碎屑盖度(%)和发达土壤上苔藓层盖度(%),都是在植被调查期间目测估计的。从实地收集的数据或从外部来源获得的每一块土地总共获得了14个环境变量(表1)。

功能性状数据:记录的植物物种的生活史特征取自国家植物区系。关于植物生长习性的数据,收集了植物生活型和开花物候的开始、结束和持续时间。此外,使用皮尔斯等人概述的方法,根据CSR生态战略对每个物种进行了表征。
数据分析:用典型对应分析(CCA)检验了不同监测样地之间植物区系组成的变化。CCA是一种单峰排序技术,其排序轴被约束为环境预测变量的线性组合。我们应用了人工正向选择程序来选择主要环境变量,从仅截取模型开始。模型的选择是基于Akaike的信息标准作为模型质量的衡量标准。最后一组约束的重要性是使用999个排列的类ANOVA排列测试来确定的。约束变量之间的相互依赖用一个方差膨胀因子来检验。物种丰度被转化为ln(x+1),并预先排除了稀有物种(仅存在于一块样地中)。// 为了提高CCA结果的可解释性,根据乔木层物种组成和母质对所有样地/立地进行了分类。首先,我们分析了每个树种在乔木层中的盖度百分比。然后对树种进行排序,并计算阔叶层和针叶层之间的比率。如果样地以单一树种为主,则被归类为单优势林分。如果乔木层有利于除青冈以外的阔叶层,则将其归入其他阔叶组。如果乔木层有利于云杉以外的其他针叶树种,则将其归入针叶树类。在该方案中,该方案包括以青冈为主的地块的地质标准,因为这些群落描绘了广泛的森林土壤和地质光谱。此外,还根据森林立地类型的植物社会学类型对每个样地进行了分类。划分出5个不同的立地类群:碳酸盐基岩上以水青冈为主的立地(12个样地)、非碳酸盐基岩上以水青冈为主的立地(8个样地)、以其他阔叶树种为主的立地(15个样地)、以云杉为主的立地(6个样地)和以其他针叶树种为主的立地(9个样地)。对于每一组,我们还通过他人提出的统计方法来确定草本层指示物种。// 使用第四角分析来评估环境变量与生活史特征之间的相关性。这一分析结合了三个矩阵:表R、表L和表Q。在社区CSR的情况下,社区加权平均值之间的关联的C分数、S分数和R分数与环境变量用一元线性回归模型检验,决定系数(R2)被用来衡量拟合优度。// 在CCA和四角分析之前,检查了环境变量的分布,并在显著偏离正态分布的情况下,使用了适当的转换。每个预测因子都被相对化,在0到1之间变化,这意味着所有的解释变量都在相同的范围内。在使用R统计软件进行的所有分析中,以P=0.05确定显著性。vegan程序包用于CCA,ade4程序包和indicspecies分别用于第四角分析和指示物种分析。

Results
植物区系组成:总体而言,最终CCA模型中包含的环境变量解释了植物群落组成变化的15.6%。最终模型中的约束变量由大到小依次为年平均温度(MAT)、年平均降水量(MAP)、乔木层遮荫能力(SCA)和土壤pH(图2)。第一和第二轴分别解释了4.6%和4.5%的变异。MAT与第一轴的相关性最强(r=-0.92),而MAP(r=0.76)、乔木层SCA(r=0.49)和土壤pH(r=0.58)与第二轴的相关性较强(图2)。

性状组成:生活史特征(特征状态)和环境变量之间共有28个关联被检测到显著。其中13个为正相关,15个为负相关(图3)。三个环境变量(年平均降水量-MAP、热负荷指数-Hi、木质碎屑盖度)不影响所调查的任何生活史特征。// 杂草生长习性受土壤pH、土壤全氮和乔木层遮荫能力(SCA)正向影响。禾本科植物与乔木层和乔木层的阔叶比例呈负相关。灌木与土壤粘粒含量和苔藓盖度呈正相关,而受乔木层特殊配合力的影响呈负相关。乔木层丰富度增加了草本层树种的频率/丰度(图3)。// 地上芽植物作为一种植物生活型,与土壤水分呈负相关。半地上部植物受土壤有机碳、土壤氮素和海拔高度的正向影响。相反,土壤中粘土的比例、乔木层物种丰富度和年平均温度对这种生活型产生了负面影响。高位芽植物表现出相反的格局。它们与粘粒比例、乔木层物种丰富度和年平均温度呈正相关,与土壤氮素和海拔呈负相关。// 在我们的数据集中,物候特征与乔木层特征联系最紧密。开花时间与乔木层和乔木层SCA中阔叶量的比例呈负相关。开花末期也是如此,与树种丰富度呈负相关。该解释变量与花期呈负相关(图3)。

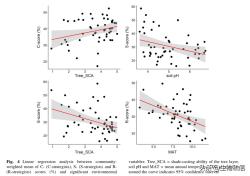
C-、S-和R-得分的社区加权平均数(CWM)与个体环境变量之间的线性回归分析显示出四个显著的关系(图4)。C分与树层SCA呈正相关。群落水平的S-值随土壤pH的升高而降低,随乔木层SCA的增加而降低。草本层群落R-S分的CWM与年均降水量呈负相关(图4)。
Discussion
这项研究强调了斯洛文尼亚温带森林草本层维管植物组合中植物区系和特征组成的环境决定因素。解释植物区系组成的主导因素。结果表明,植物区系组成受气候参数(年平均温度、年平均降水量)、土壤性质(土壤pH)和林分特征(乔木层SCA)的影响。光照条件也是匈牙利温带森林草本层组成的一个重要环境因素,光可用性驱动整个欧洲草本层群落的生物多样性变化。前人的研究证实了覆盖层对成熟森林的草本和灌木群落的有意义的影响。在我们之前的研究中,我们说明了完全没有草本层物种多样性(物种丰富度、草本层盖度和均衡度)的气候信号。结果表明,气候参数对草本层物种组成有深刻的影响。因此,植物群落结构的不同方面(组成与多样性)受到不同的环境因素的影响。除了非生物因素的强烈影响外,扩散能力、竞争或食草性等生物因素可以在更精细的空间尺度上解释森林物种分布模式。
我们方法的新颖性在于对植物区系观点和基于特征的观点进行了平行评估。尽管分类学和功能成分往往彼此紧密联系,但将重点从分类学扩大到功能特征是被广泛接受的。然而,值得注意的是,一些简单的特征(但对大多数物种来说很容易获得,例如植物的生命和生长形式)给出的一般性结论较少,因为与描述植物形态和功能的全球谱的数字特征相比,植物生态学家很少将全植物特征综合在一起。
一个值得讨论的问题是所选环境变量之间的协变性,这可能会极大地混淆对它们对物种和特征组成的实际影响的解释。例如,乔木层的组成和多样性往往与土壤属性和气候参数有关。然而,森林土壤属性也取决于主要树种、宏观气候和当地地形因素。因此,很难在不同解释因素的个体影响之间进行可靠的区分,而且直接影响可能被它们之间无数可能的相互作用所掩盖。如果森林管理或过去土地利用的遗留问题(演替发展轨迹)在构建草本层组合中发挥重要作用,预测因素之间的相互依赖可能会导致一些偏差,而准确的预测可能会更加复杂。森林群落构成是由实际营林管理直接控制的环境变量驱动的。尽管如此,基于相关矩阵(附录1),我们假设在我们的数据集中,这种混杂影响的程度相当低。同一预报组内的变量的相关性比跨预报组的高(例如,与乔木层或气候因素的关联相比,土壤参数通常显示出更高的相互关联性)。
我们选择了一组反映森林草本层植被性状变异主轴的功能性状。分类性状,如植物生命和生长形态,已被证明是解释描述叶经济学、植物大小和繁殖之间基本权衡的关键性状的变异性的良好替代品。在我们的例子中,杂草、禾本科和灌木与不同的土壤性质和乔木层特征相关。林分的这些特征受到管理干扰的影响。相同的环境因子以不同的方式影响植物的生长习性。例如,大多数森林杂草受益于茂密乔木层封育导致的较高遮荫,而禾本科植物则表现出相反的格局。改善的光照条件支持了适应强光环境的植物物种的扩展。在我们的针叶林样地中,许多草本层群落都经历了具有普遍克隆繁殖能力的草本植物的显著优势地位。这种差异对植物生命形式的影响更加明显。除土壤有机碳外,大多数与半地上部植物呈正相关的环境因子对高位芽植物都有负调控作用,反之亦然。这些模式表明,我们的数据集中排除了这两种最丰富的生命形式之间的相互作用。有报告说,地形因素(海拔和坡向)导致强烈的环境过滤,从而改变了次地中海灌木林草本层特征的相对丰富度和生命形式的功能丰富度。相反,我们的结果表明,热负荷指数(结合坡向和坡度)对任何因变量的预测都不显著。这可能是因为我们分析了斯洛文尼亚各地广泛的森林类型,而不是在较小的(区域、地方)空间尺度上比较社区。
草本层物种的开花属性(开始、结束和持续时间)仅与乔木层特征相关。结果表明,落叶林草本层群落具有开花早、持续时间短的特点。众所周知,春季地生植物适应在落叶覆盖层树冠发育之前开花,从而减少森林地面的光照供应,这反映了春季和夏季植物物种之间的物候不同步。与其他物种相比,这组物种的花期也非常短,可以根据与叶经济学和植物大小谱相关的一些关键功能性状(如比叶面积、叶干物质含量、株高、种子质量)进行额外区分。此外,林下群落的物候特征可能更多地取决于物种(即特定于物种),而不仅仅取决于郁闭度或树种组成。
CSR特征与乔木层SCA、土壤pH和年平均温度相关。乔木层SCA(光条件的代表)是最重要的因素。次地中海森林草本层特征数据的变化可以用光合作用有效辐射来最好地解释。从我们的样地模式来看,C分沿着乔木层SCA梯度增加,表明竞争对手在阴暗的林下环境中更丰富。考虑到我们40%的地块主要由山毛榉主导,这种耐荫草本植物通常出现在山毛榉林下,这可能是导致观察到的趋势的原因。山毛榉森林物种表现出相当窄的生态位范围(对土壤养分和水分的更高要求),并分享与竞争和/或抗逆策略相关的类似功能特征,但扩散能力有限。阔叶落叶欧洲森林的林下落叶层(如非酸性土壤上的山毛榉森林或峡谷森林)更常见的寄主物种具有获取叶经济)。这种模式也可以解释为草本层中的耐荫竞争树种(如青冈),这是由盖层种子来源促成的。纯山毛榉林中的草本层通常以幼树为主,主要是山毛榉和竞争草本植物。
对于大量的S-策略者来说,光线充足也是决定性的因素。然而,与通常认为森林中主要的非生物胁迫表现为低光能利用率相反,我们发现S-Score沿着乔木层SCA梯度下降。偏离预测的一种可能的解释可能是因为我们的乔木层SCA值较高的地块的特点是土壤pH值较高(这种相关性可以在图2中认识到),例如碳酸盐基岩上的不同山毛榉森林。此外,环境限制往往不会对单个性状施加选择,而是同时对多个性状施加选择。我们可以得出结论,研究数据集中的主要环境胁迫是由低土壤pH引起的,而缺乏光照似乎是一个较少的限制因素。
与低海拔森林相比,高海拔森林的自然和人为干扰(如风灾、虫害、干旱胁迫及其相关的树木死亡、雨夹雪(破冰)、抢救伐木)的影响更大。此外,云杉主要以竞争类禾本科植物为特征(例如,草类植物,如绒毛云杉或柔毛云杉),它们也表现出相当粗鲁的行为,因为它们能够在当地树冠干扰后通过广泛的营养繁殖方式传播。
总体而言,乔木层SCA被认为是与植物区系组成或性状组成(总共8个相关性)具有最高显著关联的环境因素。与林分有关的另一个重要特征是乔木层丰富度,这在5个场合被证明是显著的,其次是年平均温度(4个相关性)。在其他可能的预测因子中,树种组成、土壤pH、土壤氮素含量和土壤粘粒含量(总共表现出三个相关性)也是草本层植被组成的较强预测因子。鉴于草本层组成的主要环境决定因素与林分特征,应采取适当的森林管理干预措施,支持保护当地立地适应的乔木层物种组成、丰富度和结构,而不显著影响土层,从而保护对干扰敏感的草本植物物种。了解草本层组成和多样性的决定因素为可持续管理实践提供了理论基础。显然,保护生物多样性和促进温带森林长期可持续性的努力不能忽视林下物种。
此外,年平均温度和降水量对植物区系组成的显著影响所证明的气候信号表明,温带森林中的植物群落可能对持续的气候变化敏感,即变暖和干燥。由于功能组成在很大程度上反映了植物区系组成,因此可以预期未来气候变化对群落特征模式的类似影响。大陆尺度林下植物群落特征变异受大气候和土壤因子的驱动,受盖层盖度的调节。所有这些因素(或它们的替代者)也在我们的研究中被提及。这种分析进一步证明了评估生物多样性和功能反应的重要性,以增加我们对全球变化对草本层的可能影响的理解。气候变化是自然和人为森林干扰的导火索。基于本研究中包括的地块的子集显示,与受干扰较少的地点相比,受干扰较多的地点物种组成的时间变化更大,这表明森林干扰在十年期间导致整个斯洛文尼亚森林生态系统的组成发生重大变化。
原文链接:https://webvpn.xmu.edu.cn/https/77726476706e69737468656265737421fcfe4f976923784277068ea98a1b203a54/content/pdf/10.1007/s11258-021-01203-8.pdf








发表评论